Поперечнополосатая (скелетная) мышца — это совокупность отдельных пучков, каждый из которых насчитывает до 100 мышечных волокон (рис. 2.5, верхняя часть). В большинстве скелетных мышц каждое мышечное волокно (мышечная клетка) состоит из 1000—2000 миофибрилл. Каждая миофибрилла состоит из цепи саркомеров, последовательно соединённых «конец в конец» Основная сократительная (контрактильная) единица скелетной мышцы — это не что иное, как саркомер. Саркомеры соединены друг с другом с помощью Z-линий (или пучков), подобно связующему звену в цепях. С другой стороны, каждый саркомер содержит множество филаментов, состоящих из молекул актина и миозина, в результате взаимодействия которых и образуется сократительная (контрактильная) сила.
 В средней части рис. 2.5 показана длина саркомера в состоянии покоя мышцы вместе с полным перекрытием актиновых и миозиновых филаментов (максимальная сократительная сила). Во время максимального укорочения молекулы миозина устанавливаются напротив линии «Z», блокирующей будущее сокращение (не показано). В нижней части рис. 2.5 показано почти полное растяжение саркомера с неполным перекрыванием молекул актина и миозина (сниженная контрактильная сила).
В средней части рис. 2.5 показана длина саркомера в состоянии покоя мышцы вместе с полным перекрытием актиновых и миозиновых филаментов (максимальная сократительная сила). Во время максимального укорочения молекулы миозина устанавливаются напротив линии «Z», блокирующей будущее сокращение (не показано). В нижней части рис. 2.5 показано почти полное растяжение саркомера с неполным перекрыванием молекул актина и миозина (сниженная контрактильная сила).
Миозиновые головки миозинового филамента представляют собой определённую форму аденозинтрифосфата АТФ, которая сокращается и взаимодействует с актином, чтобы вызвать сократительную силу. Эти контакты можно наблюдать с помощью электронной микроскопии как перекрёстные мостики, расположенные между актиновыми и миозиновыми филаментами. Ионизированный кальций запускает взаимодействие между филаментами, а АТФ обеспечивает энергию. АТФ освобождает миозиновые головки от актина после одного мощного «удара» и немедленно «поднимает» его для другого цикла. Во время этого процесса АТФ превращается в аденозиндифосфат (АДФ). Ионы кальция немедленно запускают следующий цикл. Множество таких сильных «ударов» необходимо для осуществления гребневого движения, в котором задействуется множество миозиновых головок от множества филаментов, чтобы произвести одно судорожное сокращение.
В присутствии кальция и АТФ актин и миозин продолжают взаимодействовать, при этом затрагивается энергия и используется сила, чтобы сократить саркомер. Такое взаимодействие актина и миозина, в результате которого продуцируется напряжение и потребляется энергия, не может происходить, если саркомеры удлинены (мышца растянута), пока сохраняется перекрытие между актиновыми и миозиновыми головками. Это изображено в нижней части рис. 2.5, где актиновые филаменты располагаются вне пределов досягаемости половины миозиновых головок (перекрёстные мостики).
Сила сокращения, которую какому-либо саркомеру может обеспечивать напряжение при активации, зависит от его фактической длины. Сократительная сила снижается очень быстро, когда саркомер достигает максимума или минимума длины (полное растягивание или полное укорочение). Поэтому каждый саркомер мышцы может генерировать максимальную силу только в промежуточном диапазоне своей длины, но он может затрачивать энергию в состоянии полного укорочения, стараясь укоротиться ещё больше.
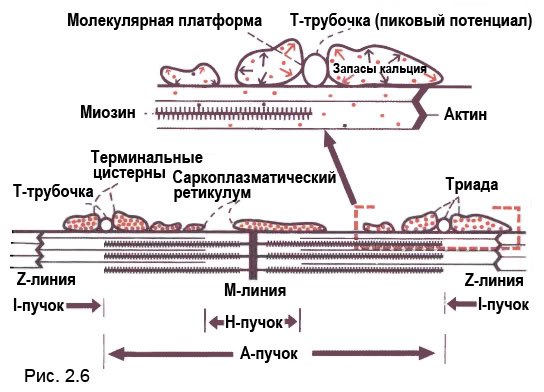
Саркоплазматический ретикулум человека состоит из трубчатой сети, которая окружает миофибриллы в мышечном волокне скелетной мышцы. Он представляет собой своеобразный резервуар кальция, который в норме высвобождается под действием пиковых потенциалов, распространяющихся вдоль поверхности мышечной клетки (сарколемма) и вдоль Т-образных трубочек (светлые круги), которые представляют собой инвагинацию сарколеммной мембраны. Изображение внизу схематически представляет один саркомер (функциональная единица скелетной мышцы), который распространяется от одной Z-линии до следующей Z-линии. Эта Z-линия находится там, где саркомеры объединяются, чтобы образовать цепь сплетающихся звеньев.
А-пучок — это область, занятая молекулами миозина (структуры, подобные щёткам), и отростками миозиновых головок.
I-пучок включает центральную Z-линию, где молекулярные филаменты актина (тонкие линии) прикрепляются к Z-линии, а I-пучок состоит из наибольшего числа филаментов. когда они свободны от перекрестных миозиновых мостиков.
М-линия образуется перекрыванием хвостиков молекулы миозина, головки которых расположены в разные стороны от М-линии.
Одна триада (две терминальные цистерны и одна Т-трубочка видны в красном квадрате) более детально показана на верхней части рисунка. Деполяризация (которая вызывается распространением типовых потенциалов вдоль Т-трубочки) передаётся через молекулярную платформу, чтобы индуцировать высвобождение кальция (красные стрелки) из саркоплазматического ретикулума. Кальций (красные точки) взаимодействует с сократительными элементами, чтобы индуцировать сократительную активность, которая продолжается до тех пор, пока кальций отсасывается внутрь саркоплазматического ретикулума или не истощатся запасы АТФ.
 В норме кальций секвестрируется в канальцевой сети capкоплазматического ретикулума (см. рис. 2.5, верхняя часть; рис. 2.6), окружающего каждую миофибриллу. Кальций высвобождается из саркоплазматического ретикулума, окружающего каждую миофибриллу, когда распространяющийся потенциал действия достигает его с поверхности клеток через «Т»-канальцы (см. рис. 2.6). В норме после высвобождения свободный кальций быстро засасывается обратно в саркоплазматический ретикулум. В отсутствие свободного кальция сократительная активность саркомеров прекращается. При отсутствии АТФ миозиновые головки остаются прочно сцепленными, а мышца становится туго напряженной, как при трупном окоченении.
В норме кальций секвестрируется в канальцевой сети capкоплазматического ретикулума (см. рис. 2.5, верхняя часть; рис. 2.6), окружающего каждую миофибриллу. Кальций высвобождается из саркоплазматического ретикулума, окружающего каждую миофибриллу, когда распространяющийся потенциал действия достигает его с поверхности клеток через «Т»-канальцы (см. рис. 2.6). В норме после высвобождения свободный кальций быстро засасывается обратно в саркоплазматический ретикулум. В отсутствие свободного кальция сократительная активность саркомеров прекращается. При отсутствии АТФ миозиновые головки остаются прочно сцепленными, а мышца становится туго напряженной, как при трупном окоченении.
Рис 2.6. Схематическое изображение одного саркомера (продольный срез), а также триады и саркоплазматического ретикулума (поперечный срез) (см. рис. 2.5 для ориентации).
Трэвелл и Симонс. "Миофасциальные боли и дисфункции"